
Ⅰ. 서 론
프로바이오틱스(probiotics)는 적정량 섭취 시 숙주의 건강에 이로움을 제공하는 살아있는 미생물로 정의되며, 장내균총 개선, 면역조절, 비만 억제, 콜레스테롤 저하, 아토피 개선 및 염증 반응 완화 등에 관여하는 것으로 보고되고 있다(Choi, 2019; Moon, 2019; Ohashi and Ushida, 2009). 최근에는 장–뇌 축(gut–brain axis)을 매개로 신경염증 조절 및 인지기능 개선 가능성이 제기되고 있으며, 장내미생물 다양성의 회복이 대사증후군 예방에 기여할 수 있다는 보고도 증가하고 있다(Le Chatelier et al., 2013; Tilg and Moschen, 2014). 이에 따라 마이크로바이옴 기반 헬스케어 산업과 개인 맞춤형 프로바이오틱스 개발이 활발히 이루어지고 있다.
그러나 생균 프로바이오틱스는 일부에서 설사, 복부 팽만감, 알레르기 반응 등의 부작용이 보고되고 있으며, 특히 면역저하자나 중환자에서 균혈증 및 패혈증 사례가 보고되면서 안전성(safety)에 대한 우려가 제기되고 있다(Kim et al., 2020; Lerner et al., 2019). 이러한 한계를 보완하기 위해 열·압력 등 물리적 처리로 불활성화된 사균체(inactivated probiotics)가 새로운 기능성 소재로 주목받고 있다(Nataraj et al., 2020; Pique et al., 2019). 사균체에는 exopolysaccharides(EPS), peptidoglycan, lipoteichoic acid, surface layer protein 등 다양한 세포벽 유래 성분이 포함되어 있으며, 이들은 장 상피세포 및 면역세포와 상호작용하여 염증 반응을 조절하고 항염 및 항산화 효과를 나타내는 것으로 보고되고 있다(Nataraj et al., 2020; Pique et al., 2019).
고령화가 진행됨에 따라 파킨슨병, 알츠하이머병, 근위축성 측삭경화증 및 다발성 경화증과 같은 퇴행성 뇌질환의 유병률이 증가하고 있다. 이들 질환의 공통적인 병리 기전으로는 활성산소종(reactive oxygen species, ROS)의 과도한 축적에 따른 산화적 스트레스와 만성 신경염증이 제시되고 있다(Oh et al., 2016; Reddy et al., 2009). 과도한 ROS는 DNA 손상, 지질과산화 및 미토콘드리아 기능 장애를 유발하고 세포사멸 신호경로를 활성화하여 신경세포 손상을 가중시킨다. 현재까지 근본적인 치료 전략은 제한적인 실정이며, 이에 따라 산화적 스트레스와 신경염증을 조절할 수 있는 기능성 소재의 개발이 요구되고 있다.
최근 프로바이오틱스 생균뿐만 아니라 사균체 등의 미생물 유래 소재가 퇴행성 뇌질환 예방 및 치료의 잠재적 대안으로 주목받고 있다. 그러나 현재까지 유산균 사균체의 신경세포 보호 활성에 대한 연구는 매우 미흡한 수준에 머물러 있으며, 이에 대한 체계적이고 광범위한 연구가 필요하다. 본 연구자들은 선행 연구를 통해 김치 유래 Lactiplantibacillus plantarum SMF398과 SMF470의 열처리 사균체가 우수한 항산화 및 항염 활성을 나타냄을 확인하였다(You et al., 2022). 이에 본 연구에서는 장–뇌축 관련 세포 간 상호작용을 간접적으로 반영한 in vitro 모델을 활용하여, H2O2 및 1-methyl-4-phenylpyridinium(MPP⁺)로 유도된 신경세포 손상 조건에서 김치 유래 L. plantarum SMF398과 SMF470 열처리 사균체의 신경보호 효과를 평가하고, 염증, 세포사멸 및 신경영양인자 관련 지표의 변화를 분석함으로써 사균체 기반 프로바이오틱스의 신경보호 가능성에 대한 기초 근거를 제시하고자 하였다.
Ⅱ. 재료 및 방법
프로바이오틱스 활성이 우수한 2종의 유산균주로는 선행연구를 통해 본 연구실에서 선별한 L. plantarum SMF398 및 L. plantarum SMF470 균주를 사용하였다. 상업용 프로바이오틱스 균주인 L. plantarum ATCC 14917은 한국생명공학연구원 생물자원센터(Korea Research Institute of Bioscience and Biotechnology, Daejeon, Korea)로부터 분양받아 사용하였다. 인간 결장 유래 세포주 HT-29(KCLB 30038)와 인간 신경모세포종 세포주 SH-SY5Y(KCLB 22266)는 한국세포주은행(Korean Cell Line Bank, Seoul, Korea)으로부터 분양받아 실험에 사용하였다.
열처리 사균체의 제조는 Cheon 등(2020)의 방법을 참고하여 수행하였다. 유산균 균주는 MRS 액체배지에서 37℃, 24h 배양한 후, phosphate-buffered saline(PBS, pH 7.2)으로 2회 세척하였다. 세척된 균체는 약 1×109 CFU/mL 농도로 조정한 후, autoclave(AC-12, JEIO TECH, Korea)를 이용하여 121℃에서 15분간 열처리하였다. 균체의 완전한 사멸 여부는 평판배양법을 통하여 확인하였다. 농도를 조정한 사균체 현탁액에 별도의 부형제는 사용하지 않았으며, 현탁액을 동결한 후 동결건조기(FDU-12000, EYELA, Korea)를 사용하여 동결건조하였다. 동결건조한 사균체는 PBS에 1 mg/mL(약 1×109 cells/mL) 농도로 재현탁하여 실험에 사용하였다.
HT-29 세포는 10% fetal bovine serum(FBS, GW Vitek, Seoul, Korea)과 1% penicillin–streptomycin solution(Cytiva, Marlborough, MA, USA)을 포함한 Roswell Park Memorial Institute(RPMI) 1640 배지(Welgene, Gyeongsan, Korea)에서 배양하였다. SH-SY5Y 세포는 10% FBS와 1% penicillin–streptomycin solution을 포함한 Dulbecco’s Modified Eagle’s Medium(DMEM, Welgene, Gyeongsan, Korea)에서 배양하였다. 두 세포주는 37℃, 5% CO2 조건이 유지되는 세포배양기(Thermo Fisher Scientific, Waltham, MA, USA)에서 배양하였다.
장–뇌축 관련 세포 간 상호작용을 간접적으로 반영하기 위해, 열처리 사균체를 HT-29 세포에 처리한 후 일정 시간 배양하고, 회수한 상등액을 여과하여 조건배지(conditioned medium, CM)를 제조하였다(Lim et al., 2020)..HT-29 세포를 6-well plate에 1×106 cells/well의 농도로 분주하고 37℃, 5% CO2 조건에서 48시간 배양한 후, 기존 배지를 제거하였다. 이후 열처리 사균체 현탁액(1×109 cells/mL) 2 mL를 각 well에 처리하여 24시간 추가 배양하였다. 배양 완료 후 상등액을 회수하여 4℃, 13,000×g에서 10분간 원심분리하였다. 분리된 상등액을 0.45 μm membrane filter로 여과하여 사균체 유래 조건배지(CM)로 사용하였다.
MTT(3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide) 분석법에 따라 SH-SY5Y 세포에 대한 조건배지의 세포독성 평가를 수행하였다(Lim et al., 2020). SH-SY5Y 세포를 96-well plate에 1×105 cells/well의 농도로 분주한 후 37℃, 5% CO2 조건에서 24시간 배양하였다. 배양 후 상등액을 제거하고, DMEM 120 μL와 조건배지 80 μL를 각 well에 첨가하여 24시간 처리하였다. 이후 세포를 PBS(pH 7.2)로 2회 세척한 다음, DMEM 180 μL와 5 mg/mL의 MTT 용액 20 μL를 첨가하여 4시간 반응시켰다. 반응 종료 후 생성된 formazan 결정을 dimethyl sulfoxide(DMSO, Daejung, Siheung, Korea) 100 μL로 용해시켰으며, microplate reader(Biochrom, Cambridge, USA)를 이용하여 570 nm에서 흡광도를 측정하였다. 대조구는 PBS(pH 7.2)를 사용하였으며, 아래의 계산식을 통해 세포생존율(%)을 산출하였다.
산화적 스트레스에 대한 조건배지의 신경세포 보호 효과는 Park 등(2017)의 방법을 참고하여 평가하였다. SH-SY5Y 세포를 96-well plate에 1×105 cells/well 농도로 분주하고 37℃, 5% CO2 조건에서 24시간 배양하였다. 배양 후 상등액을 제거하고 DMEM 100 μL와 조건배지 80 μL를 첨가하여 4시간 전처리하였다. 이후 H2O2를 20 μL 처리하여 20시간 반응시켰다. 반응 종료 후 세포를 PBS(pH 7.2)로 세척한 다음, DMEM 180 μL와 MTT 용액(5 mg/mL) 20 μL를 첨가하여 4시간 반응시켰다. 생성된 formazan 결정을 DMSO 100 μL에 용해한 후, microplate reader를 이용하여 570 nm에서 흡광도를 측정하였다. 대조구는 PBS(pH 7.2)를 사용하였으며, 아래의 계산식을 통해 세포생존율(%)을 산출하였다.
Cyclooxygenase-2(COX-2), inducible nitric oxide synthase(iNOS), Bcl-2-associated X protein(Bax), B-cell lymphoma 2(Bcl-2), brain-derived neurotrophic factor(BDNF) 유전자의 mRNA 발현 수준은 quantitative reverse transcription-PCR(qRT-PCR) 방법을 이용하여 분석하였다(Cheon et al., 2020). SH-SY5Y 세포를 6-well plate에 1×106 cells/well 농도로 분주한 후 37℃, 5% CO2 조건에서 24시간 배양하였다. 배양 후 상등액을 제거하고 DMEM 1 mL와 조건배지 800 μL 를 첨가하여 4시간 처리하였다. 이후 1 mM MPP+가 포함된 DMEM 배지 200 μL를 추가하여 20시간 반응시켰다. 반응 종료 후 PBS(pH 7.2)로 2회 세척한 다음 세포를 회수하여 RNA 추출에 사용하였다. 총 RNA는 RNeasy Mini Kit(Qiagen, Hilden, Germany)를 사용하여 제조사의 지침에 따라 추출하였으며, RNA 농도는 NanoDrop spectrophotometer(Biochrom, Cambridge, UK)를 이용하여 정량하였다. cDNA는 cDNA synthesis kit(Takara, Shiga, Japan)를 사용하여 합성하였으며, StepOnePlus™ Real-Time PCR System(Thermo Fisher Scientific, Waltham, MA, USA)을 이용하여 qRT–PCR을 수행하였다. PCR 반응 혼합물은 cDNA 2 μL, primer(Cosmogenetech, Seoul, Korea) 2 μL, nuclease-free water 6 μL, SYBR Green master mix(Cosmogenetech, Seoul, Korea) 10 μL를 포함하여 총 20 μL로 조성하였다. PCR 조건은 95℃에서 15초간 denaturation 후, 60℃에서 60초간 annealing/extension을 40회 반복하였다. GAPDH를 reference gene으로 사용하였다. 상대적 유전자 발현 수준은 delta-delta Ct(ΔΔCt)방법을 이용하여 분석하였다. PCR에 사용된 primer는 코스모진텍(Seoul, Korea)에서 구매하였으며, Table 1에 제시하였다.
| Gene | Sequence | Reference |
|---|---|---|
| BDNF | F:5′-CAAACATCCGAGGACAAGGTGG-3′ | Cheon et al., 2020 |
| R:5′-CTCATGGACATGTTTGCAGCATCT-3′ | ||
| iNOS | F:5′-CCTCGCTCTGGAAAGACCAG-3′ | Cui et al., 2020 |
| R:5′-CATGGGTTTTCCAGGCCTCT-3′ | ||
| COX-2 | F:5′-AGCATCTACGGTTTGCTGTG-3′ | Ma et al., 2019 |
| R:5′-CCTGTTTAAGCACATCGCAT-3′ | ||
| Bax | F:5′-GTGGTTGCCCTCTTCTACTTTGC-3′ | Cheon et al., 2020 |
| R:5′-GCGGACTCCAGCCACAAAGATG-3′ | ||
| Bcl-2 | F:5′-CGGCTGAAGTCTCCATTAGC-3′ | |
| R:5′-CCAGGGAAGTTCTGGTGTGT-3′ | ||
| GAPDH | F:5′-GAGTCAACGGATTTGGTCGT-3′ | Cheon et al., 2020 |
| R:5′-GACAAGCTTCCCGTTCTCAG-3′ |
모든 실험은 독립적으로 3회 반복 수행하였다. 통계분석은 GraphPad Prism 9.3(GraphPad Software, San Diego, CA, USA)을 사용하였다. 모든 결과는 평균(mean)±표준편차(standard deviation, SD)로 나타내었다. 실험군 간의 통계적 유의성은 one-way analysis of variance(ANOVA)를 이용하여 분석하였으며, 사후 검정은 Tukey’s multiple comparison test를 실시하였다. 통계적 유의성은 p<0.05를 기준으로 판단하였다.
Ⅲ. 결과 및 고찰
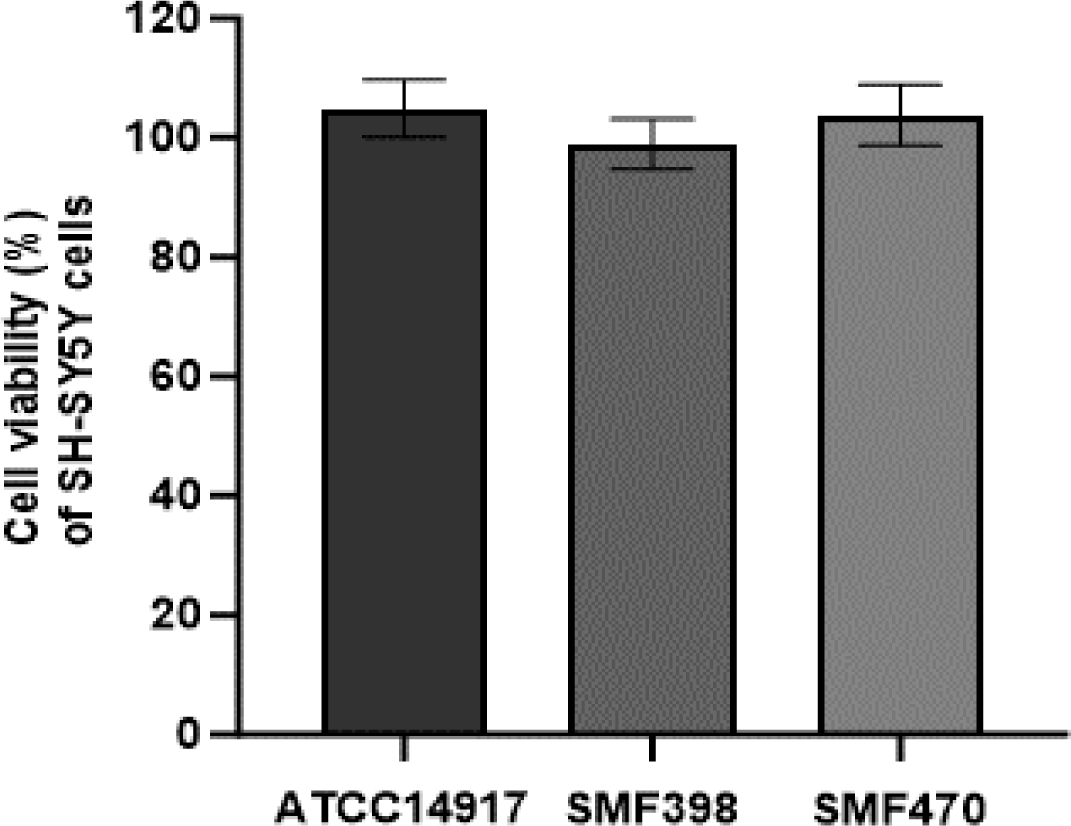
장–뇌축 관련 세포 간 상호작용을 간접적으로 반영하기 위하여, 선별된 유산균의 열처리 사균체를 인간 결장 유래 HT-29 세포에 처리하여 조건배지(conditioned medium, CM)를 제조하였다. 제조된 CM의 신경세포에 대한 세포독성은 MTT assay를 이용하여 SH-SY5Y 세포에서 평가하였다. 열처리 사균체(1 mg/mL)를 처리하여 제조한 모든 CM은 SH-SY5Y 세포에 대하여 97% 이상의 높은 생존율을 나타내었다(Fig. 1). 따라서 본 연구에서 사용된 CM은 SH-SY5Y 세포에 대해 안전한 것으로 판단되며, 신경보호 효과 평가를 위한 실험에 적합한 조건임을 확인하였다.
뇌는 높은 산소 소비율과 풍부한 다중불포화지방산 함량으로 인해 산화적 손상에 취약한 조직이며, 제한적인 항산화 방어체계로 인해 활성산소종(reactive oxygen species, ROS)이 과도하게 생성될 경우 쉽게 축적된다(Cobley et al., 2018). 이러한 ROS의 증가는 DNA 손상, 단백질 산화 및 지질과산화를 유발하여 신경세포의 기능 장애와 세포 사멸을 초래하고, 축적된 산화적 스트레스는 알츠하이머병과 파킨슨병을 포함한 퇴행성 뇌질환의 주요 병리학적 기전으로 작용하는 것으로 보고되고 있다(Uttara et al., 2009).
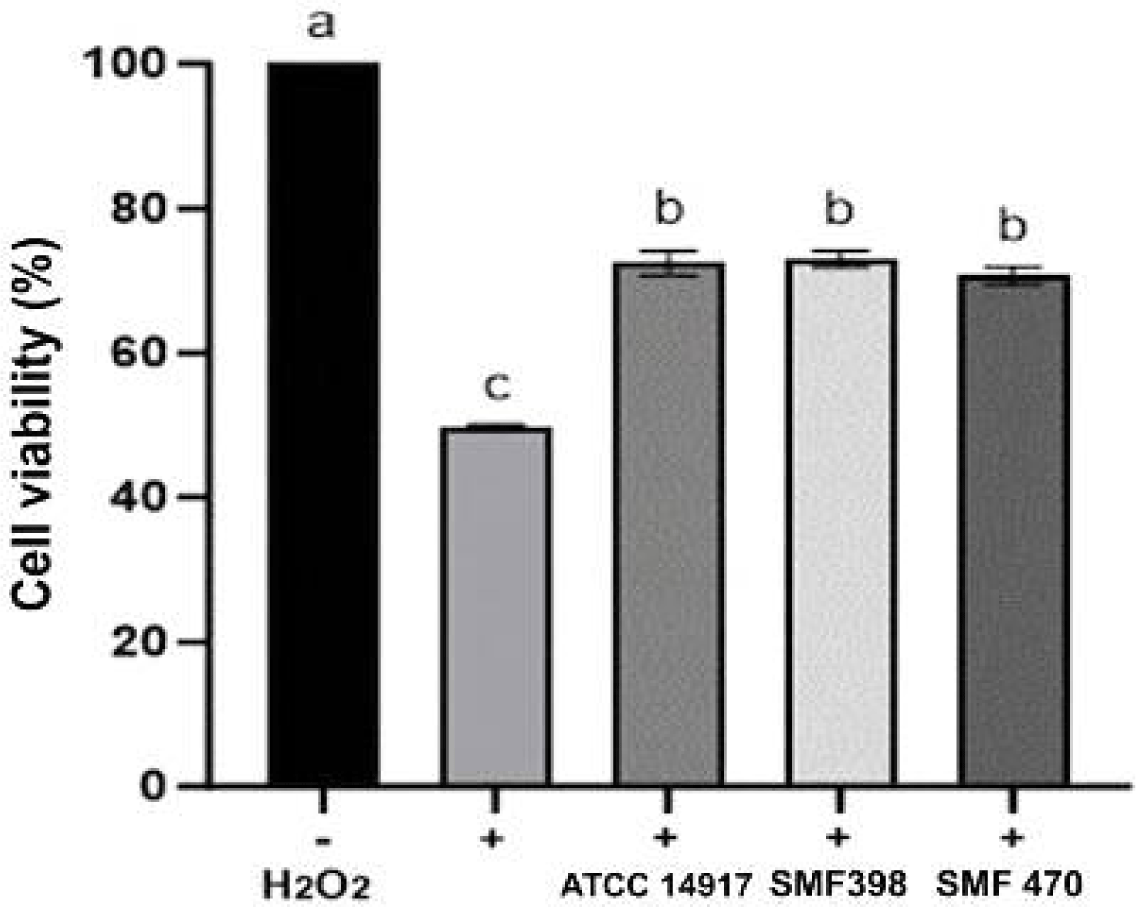
본 연구에서는 H2O2로 유도된 산화적 스트레스 조건에서 조건배지(CM)의 신경세포 보호 효과를 평가하였으며, 그 결과를 Fig. 2에 제시하였다. SH-SY5Y 세포에서 약 50%의 세포생존율을 나타내는 IC50 농도인 100 μM H2O2를 산화적 스트레스 유도 조건으로 설정하였다. H2O2 비처리군(−)은 100%의 생존율을 나타낸 반면, H2O2 처리 대조군(+)은 49.77%의 생존율을 나타내었다. 반면, H2O2로 손상된 신경세포에 SMF398 사균체 유래 CM(SMF398-CM)과 SMF470 사균체 유래 CM(SMF470-CM)을 각각 처리한 결과, 세포생존율은 72.98%와 70.65%로 나타나, H2O2 처리 대조군(+) 대비 유의적으로 생존율이 회복되었다(p<0.05). 또한 상업용 프로바이오틱스 균주인 L. plantarum ATCC 14917 유래 CM 역시 72.44%의 생존율을 보여 본 연구에서 선별된 사균체 유래 CM과 유사한 보호 효과를 나타내었다. 이러한 결과는 SMF398-CM과 SMF470-CM이 H2O2에 의해 유도된 산화적 손상으로부터 신경세포를 보호할 가능성을 시사하며, 이는 L. plantarum 사균체 유래 CM이 H2O2로 유도된 SH-SY5Y 세포의 생존율을 약 1.37배 증가시켰다고 보고한 Lim 등(2020)의 결과와 일치하는 경향을 보인다.
MPP⁺는 파킨슨병 모델에서 널리 사용되는 신경독소로, 미토콘드리아 전자전달계의 복합체 I(complex I)를 억제하여 ATP 생성을 감소시키고 ROS의 생성을 증가시킨다. 이러한 미토콘드리아 기능장애는 산화적 스트레스를 증폭시키고, 세포사멸 신호전달 경로를 활성화하여 도파민성 신경세포의 선택적 손상을 유도하는 것으로 보고되고 있다(Przedborski et al., 2003; Dias et al., 2013). 더불어 MPP⁺는 inducible nitric oxide synthase(iNOS)와 cyclooxygenase-2(COX-2)와 같은 염증성 매개인자의 발현을 증가시켜 신경염증 반응을 촉진하는 것으로 보고되었다(Cui et al., 2019; Wang et al., 2005).
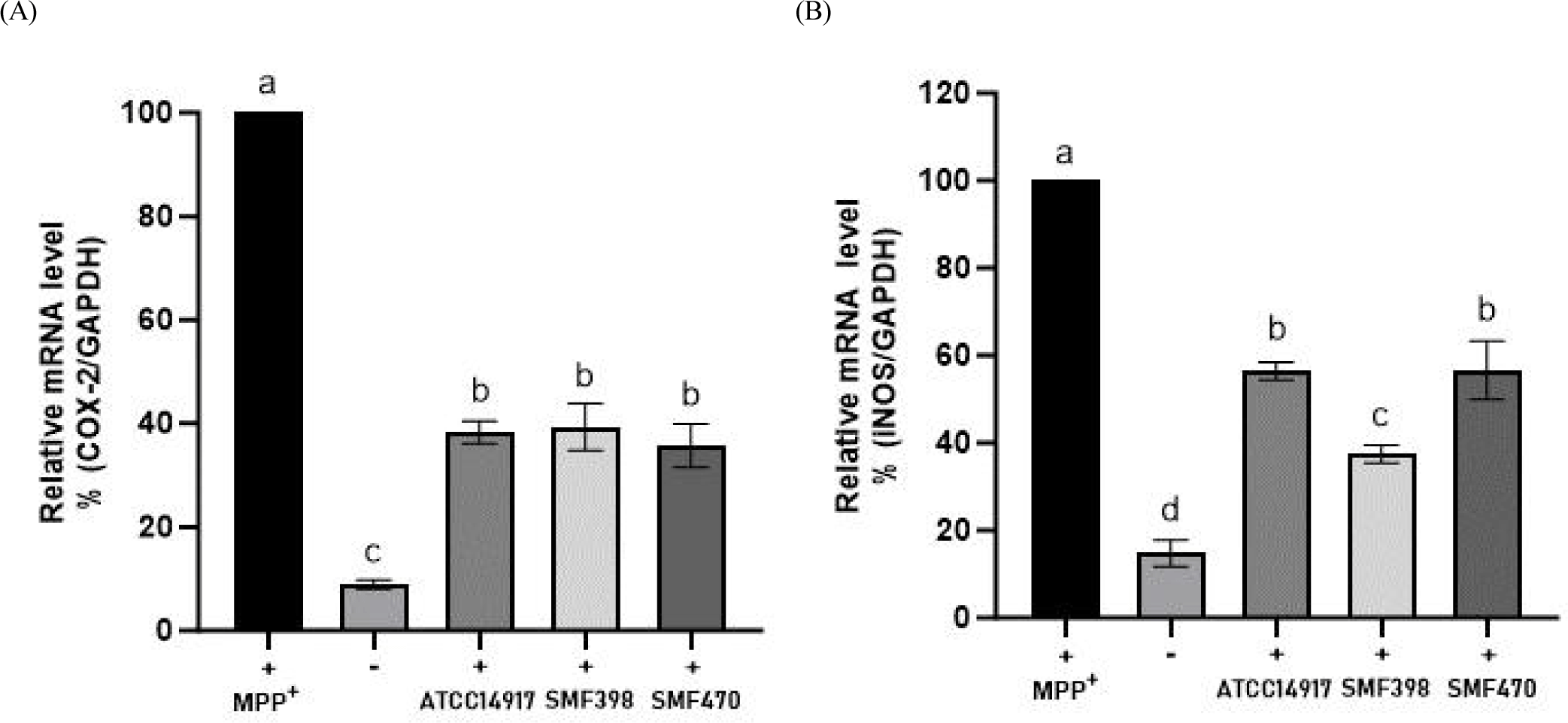
본 연구에서는 MPP⁺로 유도된 신경독성에 대한 유산균 사균체 유래 조건배지(CM)의 항염증 효과를 평가하기 위하여, MPP⁺ 처리 SH-SY5Y 세포에서 COX-2 및 iNOS의 mRNA 발현 수준을 분석하였다(Fig. 3). MPP⁺ 단독 처리 대조군(+)에서는 COX-2와 iNOS의 발현이 유의적으로 증가하였으나, CM을 처리한 군에서는 두 유전자의 발현이 모두 유의하게 감소하였다(p<0.05). 특히 SMF398-CM과 SMF470-CM 처리군에서 COX-2의 발현은 각각 60.66% 및 64.28% 감소되었으며, iNOS의 발현은 각각 62.52% 및 43.40% 감소하여 MPP⁺ 단독 처리군 대비 유의한 감소 효과를 나타내었다(p<0.05).
NO는 iNOS에 의해 생성되는 주요 염증 매개물질로, 과도하게 생성될 경우 중추신경계에서 염증 반응과 신경손상을 유발하여 파킨슨병 및 알츠하이머병과 같은 퇴행성 뇌질환의 병리와 밀접하게 관련되어 있다(Saha and Pahan, 2006). 또한 COX-2는 prostaglandin E2 합성을 촉진하는 효소로, 파킨슨병 동물모델에서 과발현되는 것으로 보고되었다(Wang et al., 2005). 따라서 본 연구 결과는 사균체 유래 CM이 MPP⁺로 유도된 염증성 유전자 발현을 억제함으로써 신경세포의 염증 반응을 조절할 가능성을 보여준다.
Bcl-2 family 단백질은 미토콘드리아 의존적 세포사멸 경로의 조절에 핵심적인 역할을 한다. Bax는 미토콘드리아 외막의 투과성을 증가시켜 cytochrome c의 방출을 유도함으로써 세포사멸을 촉진하는 반면, Bcl-2는 미토콘드리아 막을 안정화하여 세포사멸을 억제하는 항세포사멸 단백질로 작용한다(Singh et al., 2019). 따라서 Bax/Bcl-2 발현 비율은 세포사멸의 정도를 반영하는 중요한 지표로 활용된다.
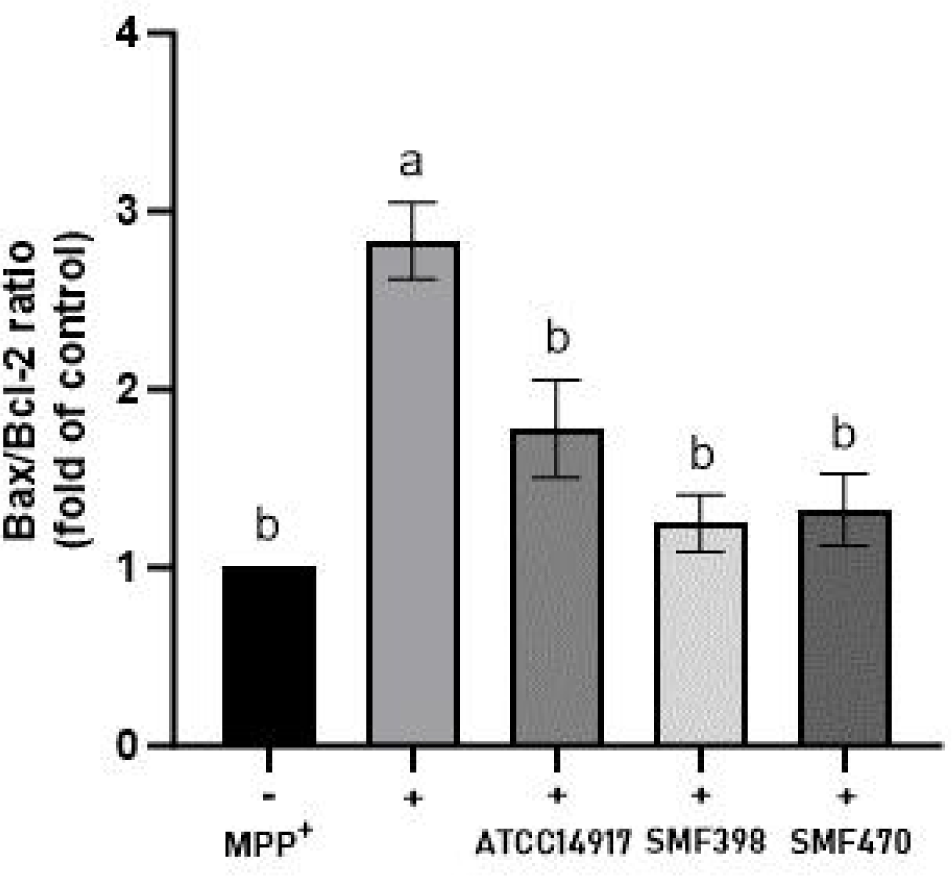
CM의 세포사멸(apoptosis) 억제 효과를 평가하기 위하여 Bax/Bcl-2 발현 비율을 분석하였다(Fig. 4). MPP⁺ 처리 대조군(+)은 비처리군(−) 대비 Bax/Bcl-2 비율이 2.83배 증가하여 MPP⁺에 의해 세포사멸이 현저히 촉진되었음을 확인하였다. 반면, SMF398-CM 및 SMF470-CM 처리군에서는 MPP⁺ 단독 처리군 대비 Bax/Bcl-2 비율이 유의적으로 감소하였다(p<0.05). Cheon 등(2020)은 MPP⁺ 처리 SH-SY5Y 세포에서 Bax/Bcl-2 비율이 대조군 대비 약 2.3배 증가하였으나, Lactobacillus buchneri 사균체 유래 CM 처리에 의해 대조군 수준으로 감소하였다고 보고하였다. 이러한 결과는 본 연구에서 관찰된 경향과 일치하며, 유산균 사균체 유래 CM이 MPP⁺로 유도된 세포사멸과 관련된 변화를 완화할 가능성을 보여준다.
CM의 신경세포 보호 효과를 추가적으로 확인하기 위하여, BDNF(brain-derived neurotrophic factor) 발현을 분석하였다(Fig. 5). BDNF는 신경세포의 생존, 분화 및 시냅스 가소성 유지에 필수적인 신경영양인자로, 신경세포 사멸을 억제하고 성장과 회복을 촉진하는 중요한 역할을 한다(Miranda et al., 2019). MPP⁺ 처리 대조군(+)에서는 BDNF 발현이 유의적으로 감소하였으나, CM 처리군에서는 발현이 유의적으로 증가되었다(p<0.05). 특히 SMF470-CM 처리군은 MPP⁺ 단독처리군 대비 약 3.1배 높은 BDNF 발현을 나타내었다. Bock 등(2023)은 H2O2로 산화적 스트레스를 유도한 SH-SY5Y 세포에서 Levilactobacillus brevis 유래 CM 처리에 의해 BDNF 발현이 증가하였다고 보고한 바 있다.
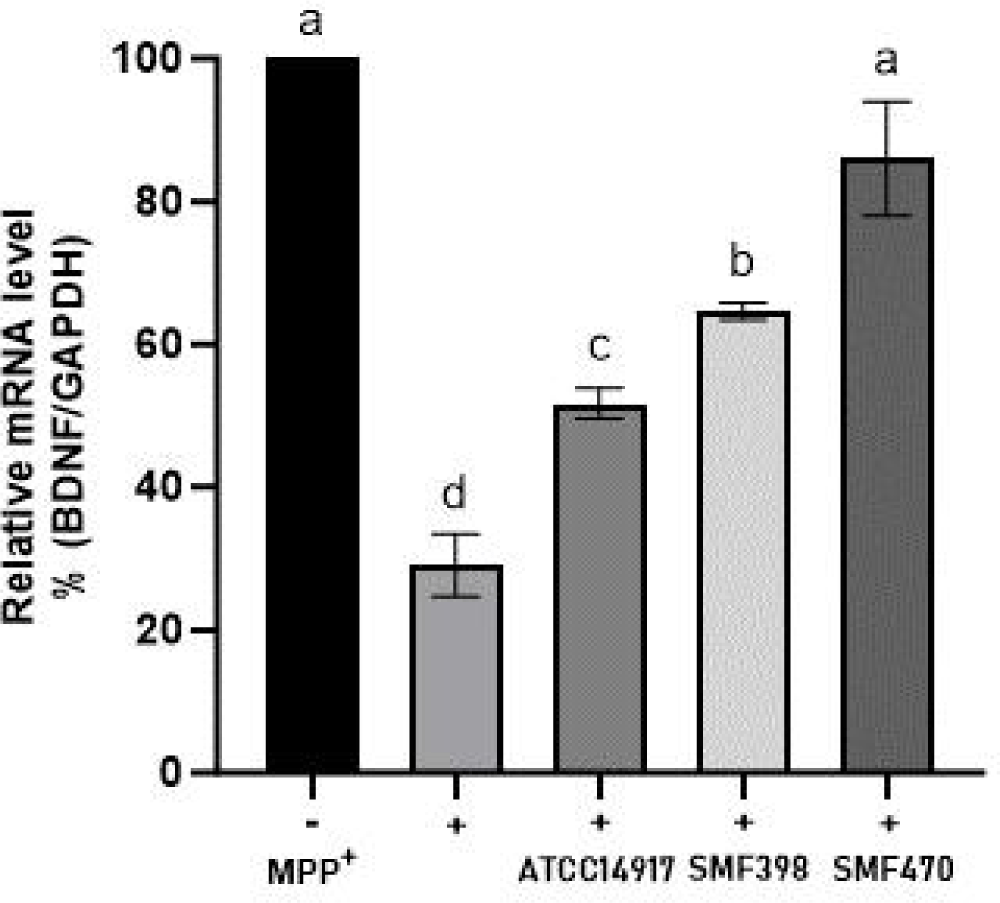
본 연구 결과, 선별된 유산균 사균체 유래 CM은 산화적 스트레스 환경에서 SH-SY5Y 세포의 생존율을 유의적으로 향상시켰으며, MPP+로 유도된 염증반응과 미토콘드리아 의존적 세포사멸을 억제하는 경향을 나타내었다. 또한 신경세포의 생존과 기능 유지에 핵심적인 역할을 하는 BDNF의 발현을 증가시킴으로써 전반적인 신경보호 효과를 나타내었다. 특히 MPP⁺로 유도된 파킨슨병 세포모델에서 염증 억제와 신경영양인자 증가가 동시에 관찰되었다는 점은, CM이 다양한 생물학적 지표의 변화와 관련되어 신경보호 효과를 나타낼 가능성을 시사한다. 이러한 효과는 유산균 세포벽 유래 성분(exopolysaccharides, peptidoglycan, lipoprotein, lipoteichoic acid, surface layer protein) 및 사균체 유래 대사산물에 의해 매개되었을 가능성이 있으며(Bock et al., 2023; Park et al., 2017; Team et al., 2020), 구체적인 작용 인자와 기전에 대해서는 향후 추가적인 연구가 필요하다. 선행연구에 따르면, 이들 성분은 염증 관련 신호전달 경로 및 산화적 스트레스 반응과 연관된 기전을 조절하는 것으로 보고되고 있으며(Nataraj et al., 2020; Pique et al., 2019), 활성산소종(ROS)의 축적은 신경퇴행성 질환의 주요 병리 기전으로 제시되고 있다(Uttara et al., 2009). 따라서 본 연구에서 관찰된 신경보호 효과 역시 이와 유사한 조절 기전과 관련되었을 가능성이 있으며, 이러한 작용은 궁극적으로 신경세포의 항상성 유지와 생존율 향상에 기여할 가능성이 있다.
Ⅳ. 요 약
본 연구는 Lactiplantibacillus plantarum SMF398 및 SMF470 열처리 사균체의 신경보호 효과를 평가하고자 수행되었다. 장–뇌축(gut–brain axis) 관련 상호작용을 간접적으로 반영하기 위해, 열처리 사균체를 처리한 HT-29 세포 유래 조건배지(conditioned medium, CM)를 SH-SY5Y 신경세포에 적용하였다. SMF398 및 SMF470 유래 CM은 H2O2로 유도된 산화적 스트레스 조건에서 SH-SY5Y 세포의 생존율을 유의적으로 향상시켰으며(p<0.05), MPP⁺로 유발된 신경독성을 완화하였다. 또한 SMF398-CM과 SMF470-CM은 MPP⁺ 처리에 의해 증가된 iNOS 및 COX-2 발현과 Bax/Bcl-2 비율을 유의적으로 감소시키고, BDNF의 발현을 유의적으로 증가시켰다(p<0.05). 이러한 결과는 김치 유래 열처리 L. plantarum 사균체가 장세포 매개 경로를 통해 신경보호 효과를 나타낼 가능성을 시사하며, 사균체 기반 기능성 소재로의 활용 가능성을 제시한다.